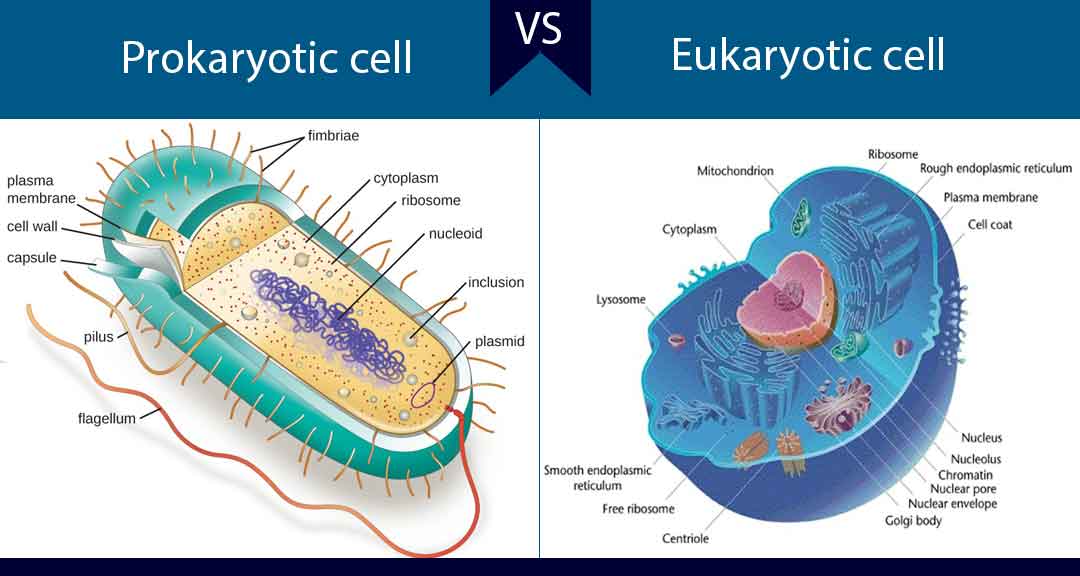
Contents
Pili are morphologically and chemically similar to fimbriae, but they are present in much smaller numbers (<10) and are usually longer. Pili are used for attachment of cells during conjugation, the process of intercellular transfer of genetic materials.
This is a semi-permeable, flexible membrane lying directly below the cell wall. It consists, principally, of a double layer of phospholipids (20 – 30%) interspersed by protein molecules (60 – 70%). It serves as an osmotic membrane for the cell, preventing the entry of undesirable solutes and allowing the absorption of essential nutrients by selective active transport.
A major difference in chemical composition between prokaryotic and eukaryotic cells is that eukaryotes have sterols in their membranes (e.g. cholesterol) whereas prokaryotes do not.
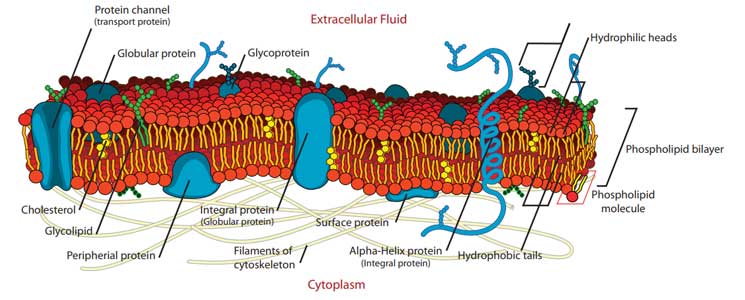
Diagram of a bacterial cytoplasmic membrane
Certain structures, known as mesosomes or chondrioids, appear in the cytoplasmic membrane. These are due to invagination of the membrane into the cytoplasm.
The mesosomes are said to be the location of several enzymes, especially the cytochrome enzyme system. In this way, the bacterial cell membrane is similar to the mitochondria found in eukaryotic cells.
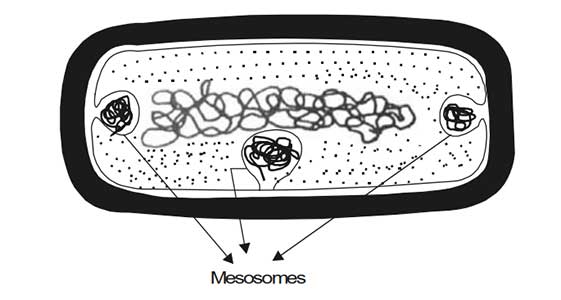
Diagrammatic structure of mesosomes
A large number of general antimicrobial agents (including surfactants) act on the cytoplasmic membrane. They cause loss of membrane integrity, leakage of intracellular contents, disruption of enzyme function, and uncoupling of proton motive force.
The bacterial cytoplasm (or simply cytoplasm) is the inner part of the cell and is enclosed directly by the cytoplasmic membrane. The cytoplasm consists of approximately 80 % water and contains enzymes that generate adenosine triphosphate (ATP) directly by oxidizing glucose and other carbon sources.
The cytoplasm also contains some of the enzymes involved in the synthesis of peptidoglycan subunits. Ribosomes, the DNA genome (nucleoid), and inclusion granules are also found in the cytoplasm.
Ribosomes are complex structures consisting of RNA and associated proteins. They are approximately 18 nm in diameter and have a molecular weight of about 2500 kDa. The ribosome is the site for protein synthesis.
Bacterial ribosomes have a sedimentation coefficient (which is a function of size) of 70s. Those found in eukaryotic cells are slightly larger and have a sedimentation coefficient of 80s. The bacterial ribosomes can dissociate into a large and a small subunit with sedimentation coefficients of 50s and 30s respectively.
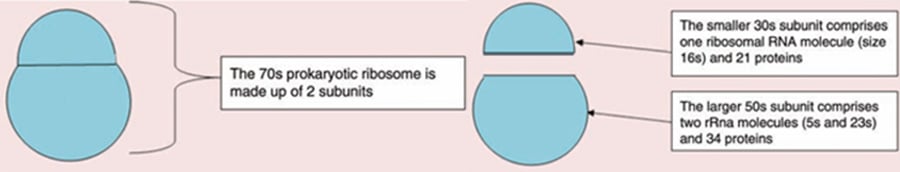
Structure of prokaryotic ribosomes
Bacterial protein synthesis starts with the interaction between a 30s ribosomal subunit, mRNA, and a tRNA (attached to the amino acid formylmethionine). The 50s ribosomal subunit then attaches and the whole ribosome moves along the mRNA chain reading the sequence of nucleotides and constructing the peptide chain dictated by the mRNA.
The nucleoid is a region of cytoplasm where the bacterial chromosomal DNA is located. The bacterial chromosome contains the genetic information necessary for functioning of the cell. It is a singular, covalently closed circular molecule of double-stranded DNA approximately 1000 times as long as the cell itself. It exists in a highly folded state and this supercoiling is brought about by topoisomerase enzymes.
Unlike in eukaryotic cells that have nuclei enclosed in a well-defined nuclear membrane, bacteria do not have a definite nuclear structure rather it exists free in the cytoplasm condensed into areas called chromatin bodies.
Bacteria often accumulate materials within storage granules within the cytoplasm. These can take a variety of forms as shown below:
Plasmids are autonomously replicating, extrachromosomal, circular pieces of double-stranded DNA. They vary in size from 1000 to 200 000 base pairs (the E. coli chromosome has 4 million base pairs) and encode for many auxiliary accessory functions conferring advantages to the cell, for example, the production of toxins, pili, bacteriocins, siderophores, and enzymes responsible for antibiotic resistance.
Plasmids replicate faster than the main bacterial genome and so cells usually contain multiple copies. All plasmids contain the information required for replication but some also contain information for cell to cell transfer (called conjugative plasmids).
Plasmids may contain multiple genes encoding antibiotic resistance elements and be able to transfer between cells. What is worrying is the ability to transfer plasmids between species, because a relatively harmless bacterium found in our gut, for example. can then donate antibiotic resistance genes to a pathogenic species, thereby increasing the spread of resistance.